The effects of distorted optic flow in multifocal glasses on self-motion perception
 Progressive addition lenses (PALs) are ophthalmic lenses mainly used to correct presbyopia. They improve far and near vision in different areas of the lens. This inevitably creates local distortions of the spatial field of view. Novice wearers of PALs often report unnatural motion percepts during self-motion. To better understand these effects, we analyzed the impact of the distorted field of view in PALs on the estimation of heading in an established computational model of self-motion perception. We simulate distorted optic flow fields for a particular PAL design and a typical self-motion scenario, a straight walk over a flat plane while fixating a point on the ground. Analyzing these flow fields showed changes in heading estimation mainly along the vertical axis. A VR-based psychophysical experiment confirmed this prediction as participants perceived upwards deviations from the true heading when gaze crosses the peripheral lens areas.
Progressive addition lenses (PALs) are ophthalmic lenses mainly used to correct presbyopia. They improve far and near vision in different areas of the lens. This inevitably creates local distortions of the spatial field of view. Novice wearers of PALs often report unnatural motion percepts during self-motion. To better understand these effects, we analyzed the impact of the distorted field of view in PALs on the estimation of heading in an established computational model of self-motion perception. We simulate distorted optic flow fields for a particular PAL design and a typical self-motion scenario, a straight walk over a flat plane while fixating a point on the ground. Analyzing these flow fields showed changes in heading estimation mainly along the vertical axis. A VR-based psychophysical experiment confirmed this prediction as participants perceived upwards deviations from the true heading when gaze crosses the peripheral lens areas.
- Y. Sauer, M. Scherff, M. Lappe, K. Rifai, N. Stein, S. Wahl. Self-motion illusions from distorted optic flow in multifocal glasses. iScience (2022), 25(1).
Motor adaptations for object size perception and grasping
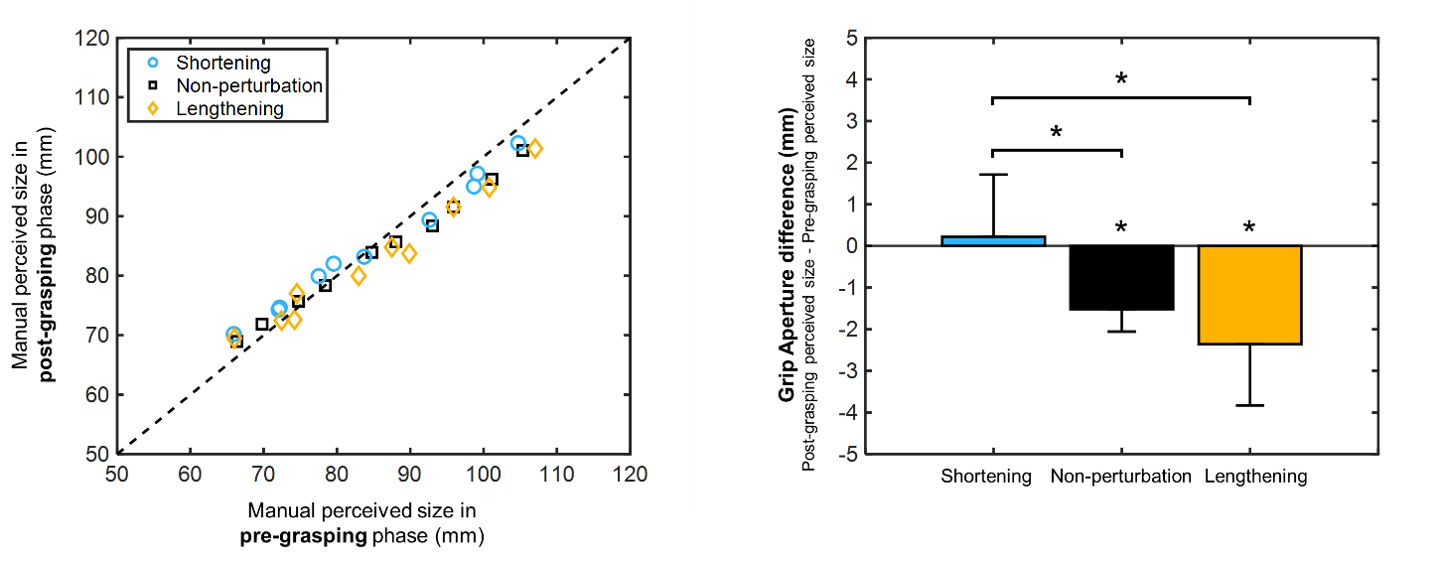 Action and perception are crucial aspects of our everyday life. Based on the dual-visual stream theory, it is now accepted that action and perception are not separated processes, but they interact and influence each other. However, it is still unknown how perception may be affected by unpredictable changes in object size during grasping movements. In this study, we aimed to determine whether size perception and saccade amplitude before and after grasping movements were modified by horizontal size perturbations during movement execution and under the presence or absence of tactile feedback.
Action and perception are crucial aspects of our everyday life. Based on the dual-visual stream theory, it is now accepted that action and perception are not separated processes, but they interact and influence each other. However, it is still unknown how perception may be affected by unpredictable changes in object size during grasping movements. In this study, we aimed to determine whether size perception and saccade amplitude before and after grasping movements were modified by horizontal size perturbations during movement execution and under the presence or absence of tactile feedback.
Two main discoveries were found. First, the horizontal size perception is modulated by the execution of grasping movements and, moreover, it relies on the presence of a horizontal size perturbation during the grasping action. Second, saccade responses exhibited amplitude modifications according to the types of target perturbations occurred in the previous action execution. These findings suggest that the combination of manual perceptual reports and corresponding saccade amplitudes are descriptive parameters of previous motor actions.
- P. Sanz Diez, A. Bosco, P. Fattori, S. Wahl. Horizontal target size perturbations during grasping movements are described by subsequent size perception and saccade amplitude. PLoS ONE 17(3): e0264560.
Effects of oscillatory optic flow on body sway
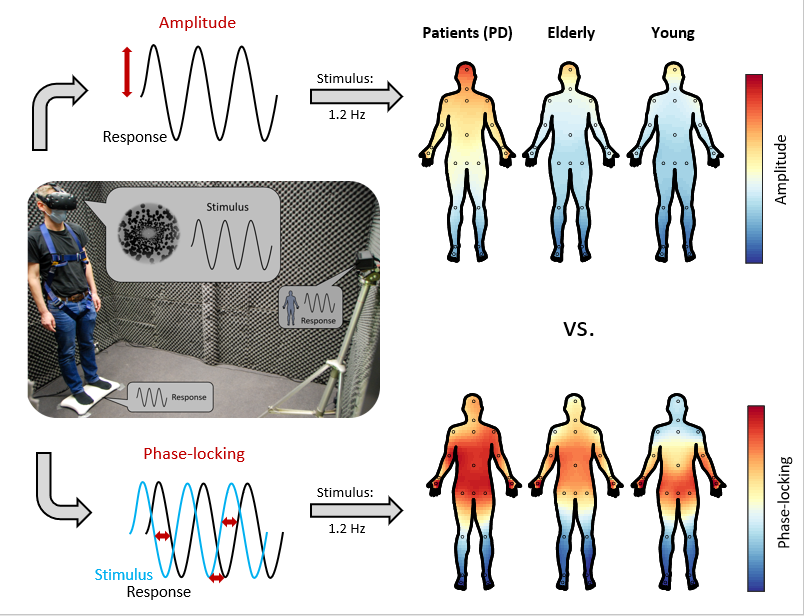 In a collaboration between UMR and MU, we established a mobile setup to investigate the impact of oscillatory optic flow on body sway and postural control. In this setup, participants stand on a force platform to track their centre of pressure while their body motion is recorded with a video-based motion tracking system. Visual stimuli are presented through a virtual reality head-mounted display.
In a collaboration between UMR and MU, we established a mobile setup to investigate the impact of oscillatory optic flow on body sway and postural control. In this setup, participants stand on a force platform to track their centre of pressure while their body motion is recorded with a video-based motion tracking system. Visual stimuli are presented through a virtual reality head-mounted display.
The innovative combination of full-body motion tracking along with our phase-locking technique, which we adopted from EEG studies and were the first to apply in the context of body sway, allowed us to reveal a previously supposed shift in participants’ movement patterns as the frequency of the visual stimulation increased.
- D. Engel, J. C. B. Schwenk, A. Schütz, A. P. Morris, F. Bremmer. Multi-segment phase coupling to oscillatory visual drive. Gait & Posture (2021) 86, 132-138.
- D. Engel, J. Student, J. C. B. Schwenk, A. P. Morris, J. Waldthaler, L. Timmermann, and F. Bremmer. Visual perturbation of balance suggests impaired motor control but intact visuomotor processing in Parkinson’s disease. Journal of Neurophysiology (2021) 126:4, 1076-1089.
Eye Tracking using real-time MRI
 We developed MREyeTrack, a novel eye tracking method based on real-time Magnetic Resonance Imaging (MRI). This allowed many fast measurements of an entire cross-section of the eye, unlike most common eye tracking that measures only the front of the eyeball. This has the advantage of measuring eye movements even when the lid is closed, additional assessment of the ocular muscles and visualising displacements and deformations of the whole eyeball. Using this new method, we could for example show that the whole eyeball is being pulled back into the eye socket during a blink. We hope that MR-based eye tracking enables new opportunities for studying ocular motility and its disorders.
We developed MREyeTrack, a novel eye tracking method based on real-time Magnetic Resonance Imaging (MRI). This allowed many fast measurements of an entire cross-section of the eye, unlike most common eye tracking that measures only the front of the eyeball. This has the advantage of measuring eye movements even when the lid is closed, additional assessment of the ocular muscles and visualising displacements and deformations of the whole eyeball. Using this new method, we could for example show that the whole eyeball is being pulled back into the eye socket during a blink. We hope that MR-based eye tracking enables new opportunities for studying ocular motility and its disorders.
- J. Kirchner, T. Watson, M. Lappe. Real-Time MRI Reveals Unique Insight into the Full Kinematics of Eye Movements. eNeuro (2021), 9(1).
Eye Tracking Latencies in Virtual Reality
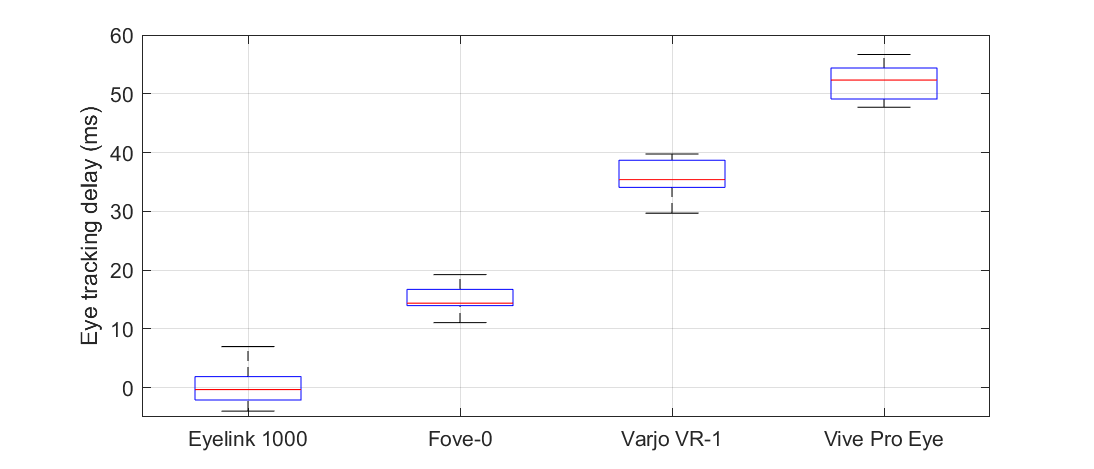
A number of virtual reality displays with integrated eye trackers have recently become commercially available. If their eye tracking latency is low and reliable enough for gaze-based rendering, this can open up many interesting opportunities for researchers. We measured eye tracking latency for the Fove-0, the Varjo VR-1, and the HTC Vive Pro Eye using simultaneous electrooculography measurements. We determined the time from the occurrence of an eye position change to its availability as data from the eye tracker (delay) and the time from an eye position change to the earliest possible change in screen content (latency). Our results show significant differences between the HMDs. Delays ranged from 15 ms to 52 ms, and latencies ranged from 45 ms to 81 ms. The Fove-0 seems to be the fastest device and is best suited for gaze-based rendering.
- N. Stein, D. C. Niehorster, T. Watson, F. Steinicke, K. Rifai, S. Wahl, M. Lappe. A Comparison of Eye Tracking Latencies Among Several Commercial Head-Mounted Displays. i-Perception (2021).
Adaptation to geometrically skewed moving images: An asymmetrical effect on the Double-Drift Illusion
 We used the curveball illusion to better understand how motion is perceived under the effect of skew distortions in our peripheral visual field. This illusion is a drifting texture object that, while falling straight in the periphery, fools our perception into believing its trajectory is tilted in the drift direction.
We used the curveball illusion to better understand how motion is perceived under the effect of skew distortions in our peripheral visual field. This illusion is a drifting texture object that, while falling straight in the periphery, fools our perception into believing its trajectory is tilted in the drift direction.
After a short period of watching a skew distorted movie, we observed that motion and the illusory perception of motion get affected in a particular way. We prove that the tuned cells by skew distortions affect complex perceptual effects, like illusory perception. Progressive addition lenses that induce this kind of distortions in the periphery may change how we usually interact with the perceived motion and become a reason for discomfort.
M. García García, K. Rifai, S. Wahl & T. Watson (2021). Adaptation to geometrically skewed moving images: An asymmetrical effect on the double-drift illusion. Vision Research 2021, 179 (75-84).
Anatomic investigation of the Parietal Reach Region
 The medial bank of the intraparietal sulcus encompasses a parietal reach region (PRR) where neurons are involved in the planning of visually guided arm movements, which has been the subject of interest in research related to prosthetic arm control. To clarify the anatomic subdivisions of PRR, we examined the connections of different sites within and rostral to this region with other areas of the macaque cortex. Based on differences in the density and modality specificity of connections and histologic characteristics, we propose a subdivision of the medial bank into areas.
The medial bank of the intraparietal sulcus encompasses a parietal reach region (PRR) where neurons are involved in the planning of visually guided arm movements, which has been the subject of interest in research related to prosthetic arm control. To clarify the anatomic subdivisions of PRR, we examined the connections of different sites within and rostral to this region with other areas of the macaque cortex. Based on differences in the density and modality specificity of connections and histologic characteristics, we propose a subdivision of the medial bank into areas.
This anatomic scheme, which incorporates features of previous proposals but refines the boundaries, may help guide future studies aimed at clarifying the functions of different medial intraparietal areas.
- S. Bakola, L. Passarelli, T. Huynh, D. Impieri, K. H. Worthy, P. Fattori, C. Galletti, K. J. Burman, M. G. P. Rosa. Cortical Afferents and Myeloarchitecture Distinguish the Medial Intraparietal Area (MIP) from Neighboring Subdivisions of the Macaque Cortex. eneuro
Size perception during saccades
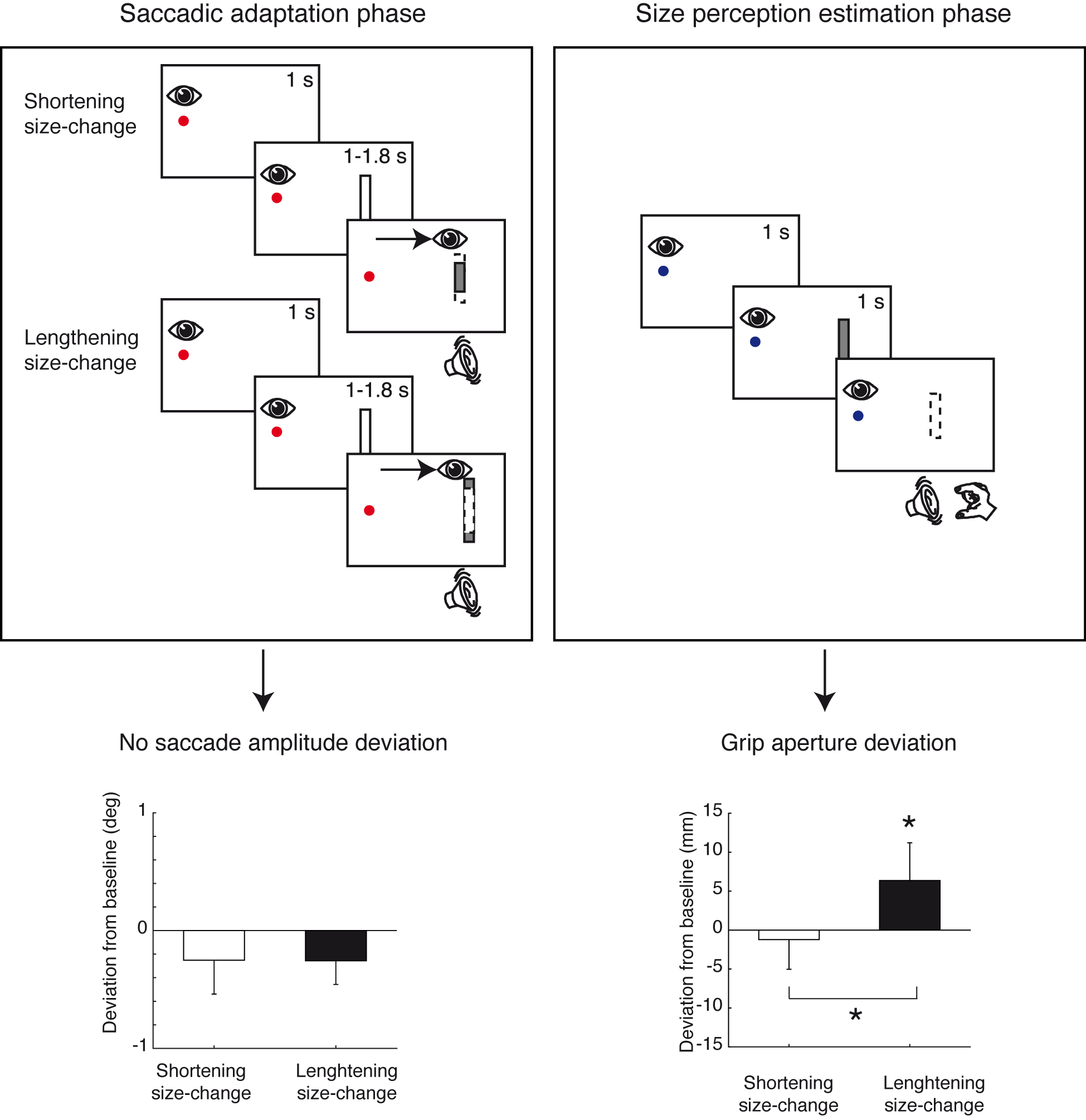 Systematic shortening or lengthening of target objects during saccades modifies saccade amplitudes and perceived size of the objects.
Systematic shortening or lengthening of target objects during saccades modifies saccade amplitudes and perceived size of the objects.
In the present study, we asked whether or not the two are necessarily linked. We tested human participants in symmetrical systematic shortening and lengthening of a vertical bar during a horizontal saccade, aiming to not modify the saccade amplitude.
By this manipulation, we did not induce any change in horizontal or vertical components of saccade amplitude, but we found a significant difference in perceived size after the lengthening experiment compared to after the shortening experiment suggesting mechanisms not depending on saccadic amplitude change.
- A. Bosco, R. Breveglieri, M. Filippini, C. Galletti, P. Fattori. Reduced neural representation of arm/hand actions in the medial posterior parietal cortex. Sci Rep 9, 936 (2019).
- A. Bosco, K. Rifai, S. Wahl, P. Fattori, M. Lappe. Trans-saccadic adaptation of perceived size independent of saccadic adaptation. Journal of Vision 2020;20(7):19.
Adaptation to Increased Curvature

Walking is the most natural way to move in virtual reality, but the limited physical space for movement restricts its applicability. One option is to alter the user’s view of the virtual scene to steer them along a specific path and give them the impression that they are moving in a straight line in a large virtual space. We tested whether prolonged exposure to this manipulation would alter perceptual manipulation thresholds. Therefore, participants first completed a measurement of their perceptual threshold. In a second session, the same participants were exposed to a virtual environment with a substantial curvature gain in which they walked between two targets for approximately 20 minutes. Afterwards, their perceptual thresholds were measured again . The results showed that thresholds increased.
- L. Bölling, N. Stein, F. Steinicke and M. Lappe. Shrinking Circles: Adaptation to Increased Curvature Gain in Redirected Walking. in IEEE Transactions on Visualization and Computer Graphics, vol. 25, no. 5, pp. 2032-2039, May 2019.
Control of saccadic adaptation
 We studied inhibition of saccadic adaptation in an eye tracking study. We wondered if saccadic adaptation is accessible to volitional control, specifically whether any adaptive change in saccade amplitude can be inhibited. Participants were repeatedly exposed to post-saccadic error using the double-step paradigm, in which the saccade target object is shifted during the eye movement. In one condition, participants were instructed to follow the target step and look at the final target location. In the other condition, participants were instructed to inhibit the adjustment of saccade amplitude and to look at the initial target location despite possible intra-saccadic movement of the target. We found that when participants were instructed to inhibit, there were no significant adaptive changes in saccade amplitude following outward target steps, but there was residual adaptation for inward target shifts. The findings indicate that volitional control can be exerted on adaptation of saccade amplitude.
We studied inhibition of saccadic adaptation in an eye tracking study. We wondered if saccadic adaptation is accessible to volitional control, specifically whether any adaptive change in saccade amplitude can be inhibited. Participants were repeatedly exposed to post-saccadic error using the double-step paradigm, in which the saccade target object is shifted during the eye movement. In one condition, participants were instructed to follow the target step and look at the final target location. In the other condition, participants were instructed to inhibit the adjustment of saccade amplitude and to look at the initial target location despite possible intra-saccadic movement of the target. We found that when participants were instructed to inhibit, there were no significant adaptive changes in saccade amplitude following outward target steps, but there was residual adaptation for inward target shifts. The findings indicate that volitional control can be exerted on adaptation of saccade amplitude.
- F. Heins, A. Meermeier, M. Lappe. Volitional control of saccadic adaptation. PLoS ONE (2019) 14(1): e0210020.
Decoding of reach actions in the Parietal Cortex
 Depths and directions of reaching actions are reliably decoded by the parietal cortex
Depths and directions of reaching actions are reliably decoded by the parietal cortex
Goal locations are discriminated well before movement onset
The entire prehension action could be decoded from a single cortical site
Neuroprosthetics from the medial parietal cortex can restore lost reach/grasp functions
- M. Filippini, R. Breveglieri, K. Hadjidimitrakis, A. Bosco, P. Fattori. Prediction of Reach Goals in Depth and Direction from the Parietal Cortex. Cell Reports 23(3), 725-731.
Reach movement encoding in macaque Area V6A
 The medial posterior parietal cortex is involved in the encoding of reaching and grasping actions in different visual conditions mainly investigated at single-cell level.
The medial posterior parietal cortex is involved in the encoding of reaching and grasping actions in different visual conditions mainly investigated at single-cell level.
In the present study, we looked for a “low-dimensional” representation of these encoding processes by studying macaque V6A neurons tested in three different tasks with a dimensionality reduction technique, which is very suitable for neuroprosthetics readout.
Comparing the portions of population variance involved in the encoding of visual information, target position, wrist orientation and grip type, we found a different use of this task relevant information according to the type of planned and executed action.
- M. De Vitis, R. Breveglieri, K. Hadjidimitrakis, W. Vanduffel, C. Galletti, P. Fattori. The neglected medial part of macaque area PE: segregated processing of reach depth and direction. Brain Structure & Function (2019), 224(7), pp. 2537 – 2557.
Uniform coding of space in the parietal cortex
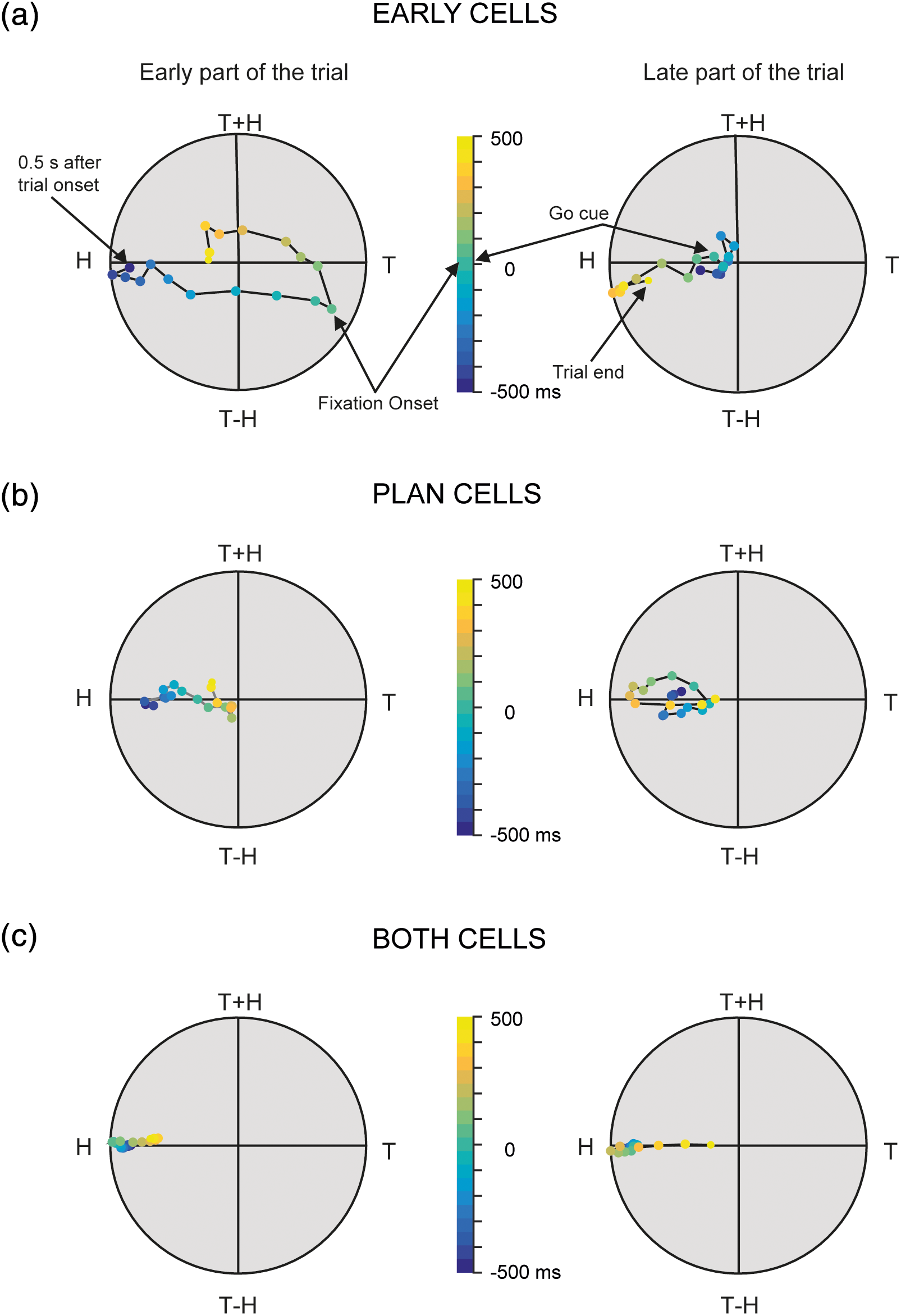 How our brain encodes the spatial location of objects in our environment and uses this information to control goal-directed movements like reaching and grasping?
How our brain encodes the spatial location of objects in our environment and uses this information to control goal-directed movements like reaching and grasping?
Despite numerous studies, there is no consensus on the degree of stability in the spatial representations across the rich repertoire of manual actions. Hadjidimitrakis et al. studied the spatial reference frames of single cells in parietal cortex and compared them between two different types of motor response evoked by the same stimulus. Parietal cortex activity showed similar reference frames across the two tasks suggesting a uniform coding of space that enables flexible motor control.
- K. Hadjidimitrakis, S. Bakola, T. Wong Yan, A. Hagan Maureen. Mixed Spatial and Movement Representations in the Primate Posterior Parietal Cortex. Frontiers in Neural Circuits 13 (2019).
- K. Hadjidimitrakis, S. Bakola, T. A. Chaplin, Hsin-Hao Yu, O. Alanazi, J. M. Chan, K. H. Worthy, M. G. P. Rosa. Topographic Organization of the ‘Third-Tier’ Dorsomedial Visual Cortex in the Macaque. Journal of Neuroscience (2019), 39 (27) 5311-5325.
Connectivity of the superior parietal lobule
 The subcortical inputs from the thalamus are in agreement with cortico-cortical circuits and indeed areas of the SPL receive from lateral posterior and medial pulvinar nuclei that integrates visuo-motor information, motor functions from the ventral lateral nucleus and eye-related activity from medial dorsal nucleus.
The subcortical inputs from the thalamus are in agreement with cortico-cortical circuits and indeed areas of the SPL receive from lateral posterior and medial pulvinar nuclei that integrates visuo-motor information, motor functions from the ventral lateral nucleus and eye-related activity from medial dorsal nucleus.
- M. Gamberini, L. Passarelli, P. Fattori, P., C. Galletti, Structural connectivity and functional properties of the macaque superior parietal lobule. Brain Struct Funct (2020) 225, 1349–1367.
- L. Passarelli, M. Gamberini, P. Fattori. The superior parietal lobule of primates: a sensory-motor hub for interaction with the environment. J. Integr. Neurosci. 2021, 20(1), 157–171.
Functional propertys of the superior parietal lobule
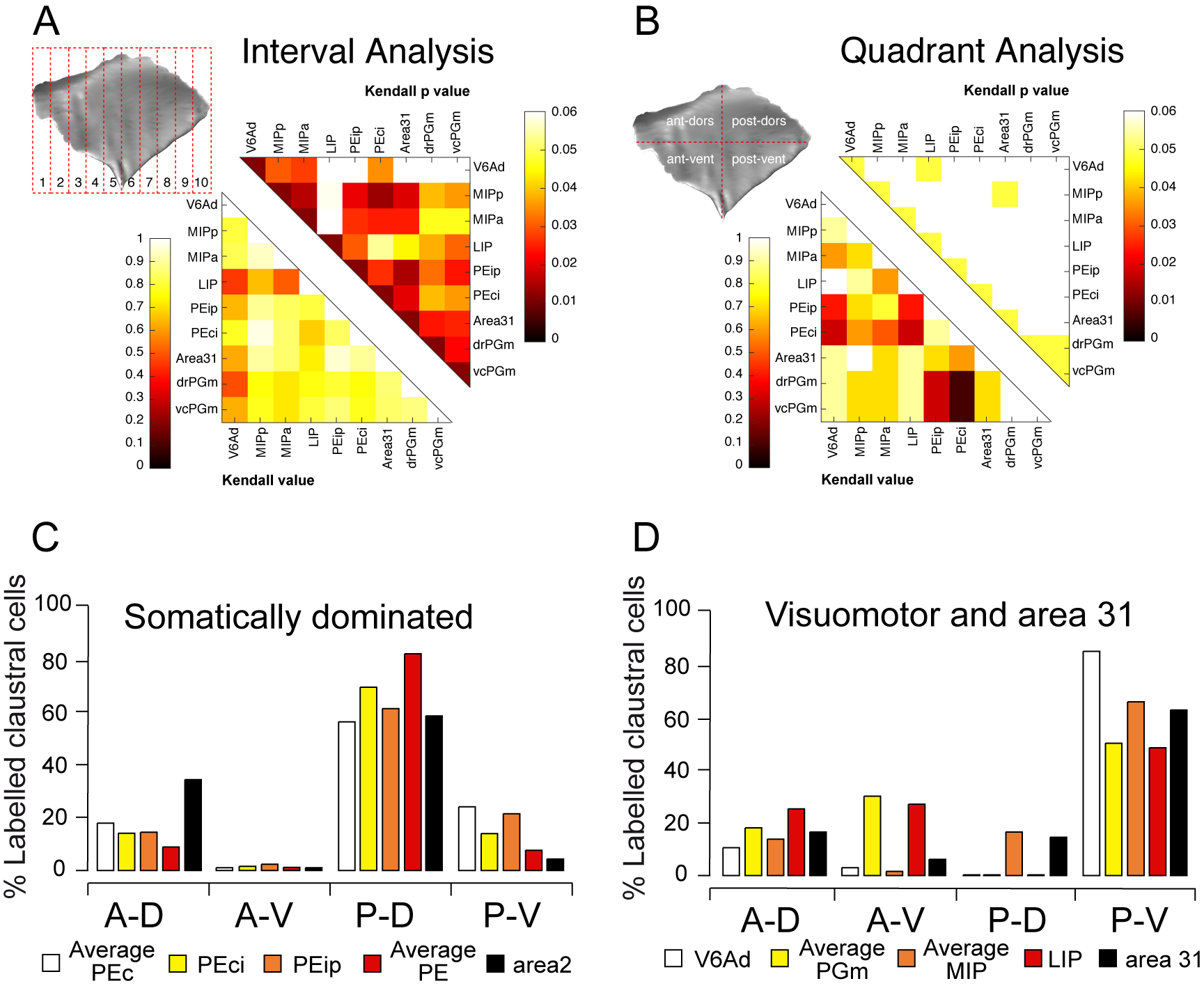 All the areas of SPL are directly connected with the claustrum. The claustral connections well agree with the functional characteristics of SPL areas and, even if a certain level of variability is evident, they receive inputs mainly from the posterior portion of this subcortical structure known to be involved in sensory encoding (visual and somatosensory).
All the areas of SPL are directly connected with the claustrum. The claustral connections well agree with the functional characteristics of SPL areas and, even if a certain level of variability is evident, they receive inputs mainly from the posterior portion of this subcortical structure known to be involved in sensory encoding (visual and somatosensory).
- M. Gamberini, L. Passarelli, P. Fattori, C. Galletti. Structural connectivity and functional properties of the macaque superior parietal lobule. Brain Struct Funct (2020) 225, 1349–1367.
Superior parietal lobule in humans
 In macaques, the superior parietal lobule (SPL) integrates information from various cortical sectors like visual (V) motor (M) and somatosensory (S) cortex and from the thalamus (TH) and thus forms an interface between perception and action. Recent functional imaging studies have identified putative human homologues of several macaque SPL areas.
In macaques, the superior parietal lobule (SPL) integrates information from various cortical sectors like visual (V) motor (M) and somatosensory (S) cortex and from the thalamus (TH) and thus forms an interface between perception and action. Recent functional imaging studies have identified putative human homologues of several macaque SPL areas.
Knowledge of this part of the brain could help understanding abnormal reaching and grasping behavior in humans and also fuel research on brain machine interfaces (BMI) enabling prosthetic arms and hand to perform more smooth and accurate actions In dynamic environments.
- L. Passarelli, M. Gamberini, P. Fattori. The superior parietal lobule of primates: a sensory-motor hub for interaction with the environment. J. Integr. Neurosci. 2021, 20(1), 157–171.
Neuronal response to visually simulated self-motion
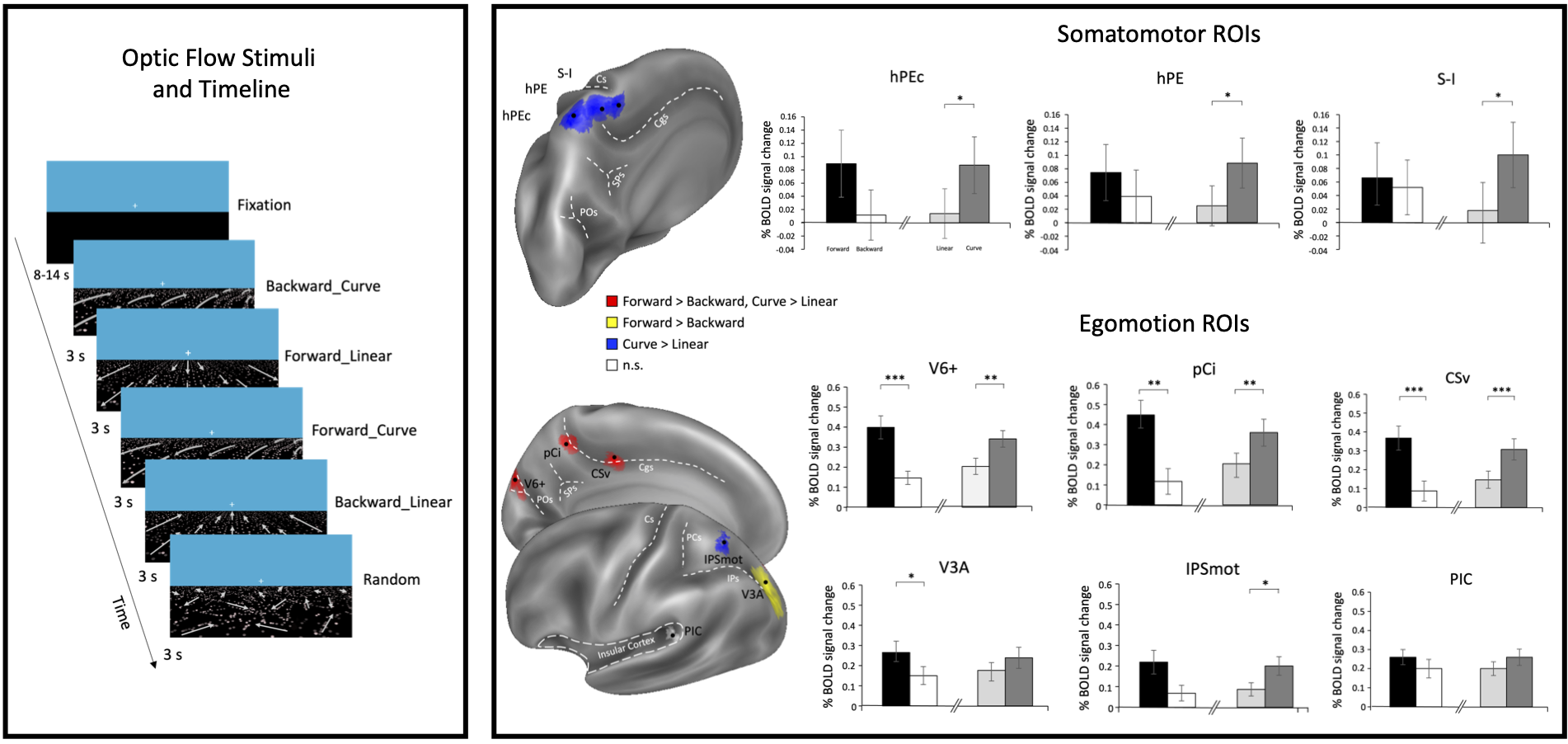 In an fMRI study the neural response to visual aspects of the optic flow generated during locomotion (passive viewing of stimuli simulating linear/curved paths and forward/backward direction of movement) has been investigated in egomotion- (V6+, V3A, IPSmot, CSv, pCi, PIC) and somatomotor-related (hPEc, hPE, S-I) cortical regions of interest (ROIs), likely involved in the visual processing aimed at locomotion control.
In an fMRI study the neural response to visual aspects of the optic flow generated during locomotion (passive viewing of stimuli simulating linear/curved paths and forward/backward direction of movement) has been investigated in egomotion- (V6+, V3A, IPSmot, CSv, pCi, PIC) and somatomotor-related (hPEc, hPE, S-I) cortical regions of interest (ROIs), likely involved in the visual processing aimed at locomotion control.
ROIs exhibited differential preference for curved paths (for controlling changing direction of movement) and/or forward direction (for a fine visual analysis of the environment needed during locomotion) suggesting that ROIs play a differentiated role in the visual guidance of locomotion.
- S. Di Marco, P. Fattori, G. Galati, C. Galletti, M. Lappe, T. Maltempo, C. Serra, V. Sulpizio, S. Pitzalis. Preference for locomotion-compatible curved paths and forward direction of self-motion in somatomotor and visual areas. Cortex (2021), 137 74-92.
List of publications from Platypus (open access)
- S. Bakola, L. Passarelli, T. Huynh, D. Impieri, K. H. Worthy, P. Fattori, C. Galletti, K. J. Burman, M. G. P. Rosa. Cortical Afferents and Myeloarchitecture Distinguish the Medial Intraparietal Area (MIP) from Neighboring Subdivisions of the Macaque Cortex. eneuro. doi: 10.1523/ENEURO.0344-17.2017
- L. Bölling, N. Stein, F. Steinicke and M. Lappe. Shrinking Circles: Adaptation to Increased Curvature Gain in Redirected Walking. in IEEE Transactions on Visualization and Computer Graphics, vol. 25, no. 5, pp. 2032-2039, May 2019. doi: 10.1109/TVCG.2019.2899228
- A. Bosco, K. Rifai, S. Wahl, P. Fattori, M. Lappe. Trans-saccadic adaptation of perceived size independent of saccadic adaptation. Journal of Vision 2020;20(7):19. doi: 10.1167/jov.20.7.19
- A. Bosco, R. Breveglieri, M. Filippini, C. Galletti, P. Fattori. Reduced neural representation of arm/hand actions in the medial posterior parietal cortex. Sci Rep 9, 936 (2019). doi: 10.1038/s41598-018-37302-2
- D. Engel, A. Schütz, M. Krala, J. C. B. Schwenk, A. Morris, F. Bremmer. Inter-trial phase coherence of visually evoked postural responses in virtual reality. Exp Brain Res 238, 1177–1189 (2020). doi: 10.1007/s00221-020-05782-2
- D. Engel, J. Student, J. C. B. Schwenk, A. P. Morris, J. Waldthaler, L. Timmermann, and F. Bremmer. Visual perturbation of balance suggests impaired motor control but intact visuomotor processing in Parkinson’s disease. Journal of Neurophysiology (2021) 126:4, 1076-1089. doi: 10.1152/jn.00183.2021
- D. Engel, J. C. B. Schwenk, A. Schütz, A. P. Morris, F. Bremmer. Multi-segment phase coupling to oscillatory visual drive. Gait & Posture (2021) 86, 132-138. doi: 10.1016/j.gaitpost.2021.03.010
- M. Filippini, R. Breveglieri, K. Hadjidimitrakis, A. Bosco, P. Fattori. Prediction of Reach Goals in Depth and Direction from the Parietal Cortex. Cell Reports 23(3), 725-731. doi: 10.1016/j.celrep.2018.03.090
- M. Gamberini, L. Passarelli, P. Fattori, C. Galletti. Structural connectivity and functional properties of the macaque superior parietal lobule. Brain Struct Funct (2020) 225, 1349–1367. doi: 10.1007/s00429-019-01976-9
- M. Garcia Garcia, K. Rifai, S. Wahl, T. Watson. Adaptation to geometrially skewed moving images: An asymmetrical effect on the double-drift illusion. Vision Research (2021), 179 75-84. doi: 10.1016/j.visres.2020.11.008
- S. Gremmler, M. Lappe. Postsaccadic eye position contributes to oculomotor error estimation in saccadic adaptation. Journal of Neurophysiology (2019) 122:5, 1909-1917. doi: 10.1152/jn.00095.2019
- S. W. Habtegiorgis, K. Rifai, M. Lappe, S. Wahl. Experience-dependent long-term facilitation of skew adaptation. Journal of Vision 2018;18(9):7. doi: 10.1167/18.9.7.a
- K. Hadjidimitrakis. Coupling of head and hand movements during eye-head-hand coordination: there is more to reaching than meets eye. Journal of Neurophysiology (2020) 123:5, 1579-1582. doi: 10.1152/jn.00099.2020
- K. Hadjidimitrakis, S. Bakola, T. Wong Yan, A. Hagan Maureen. Mixed Spatial and Movement Representations in the Primate Posterior Parietal Cortex. Frontiers in Neural Circuits 13 (2019). doi: 10.3389/fncir.2019.00015
- K. Hadjidimitrakis, S. Bakola, T. A. Chaplin, Hsin-Hao Yu, O. Alanazi, J. M. Chan, K. H. Worthy, M. G. P. Rosa. Topographic Organization of the ‘Third-Tier’ Dorsomedial Visual Cortex in the Macaque. Journal of Neuroscience (2019), 39 (27) 5311-5325. doi: 10.1523/JNEUROSCI.0085-19.2019
- F. Heins, A. Meermeier, M. Lappe. Volitional control of saccadic adaptation. PLoS ONE (2019) 14(1): e0210020. doi: 10.1371/journal.pone.0210020
- J. Kirchner, T. Watson, M. Lappe. Real-Time MRI Reveals Unique Insight into the Full Kinematics of Eye Movements. eNeuro (2021), 9 (1). doi: 10.1523/ENEURO.0357-21.2021
- R. Lencer, I. Meyhöfer, J. Triebsch, K. Rolfes, M. Lappe, T. Watson. Saccadic suppression in schizophrenia. Sci Rep 11, 13133 (2021). doi: 10.1038/s41598-021-92531-2
- S. Di Marco, P. Fattori, G. Galati, C. Galletti, M. Lappe, T. Maltempo, C. Serra, V. Sulpizio, S. Pitzalis. Preference for locomotion-compatible curved paths and forward direction of self-motion in somatomotor and visual areas. Cortex (2021), 137 74-9. doi: 10.1016/j.cortex.2020.12.021
- L. Passarelli, M. Gamberini, P. Fattori. The superior parietal lobule of primates: a sensory-motor hub for interaction with the environment. J. Integr. Neurosci. 2021, 20(1), 157–171. doi: 10.31083/j.jin.2021.01.334
- P. Sanz Diez, A. Bosco, P. Fattori, S. Wahl. Horizontal target size perturbations during grasping movements are described by subsequent size perception and saccade amplitude. PLoS ONE 17(3): e0264560. doi: 10.1371/journal.pone.0264560
- Y. Sauer, M. Scherff, M. Lappe, K. Rifai, N. Stein, S. Wahl. Self-motion illusions from distorted optic flow in multifocal glasses. iScience (2022), 25(1). doi: 10.1016/j.isci.2021.103567
- J. C. B. Schwenk, R. VanRullen, F. Bremmer. Dynamics of Visual Perceptual Echoes Following Short-Term Visual Deprivation. Cerebral Cortex Communications, Volume 1, Issue 1, 2020, tgaa012. doi: 10.1093/texcom/tgaa012
- N. Stein, D. C. Niehorster, T. Watson, F. Steinicke, K. Rifai, S. Wahl, M. Lappe. A Comparison of Eye Tracking Latencies Among Several Commercial Head-Mounted Displays. i-Perception (2021). doi: 10.1177/2041669520983338
- J. Student, D. Engel, L. Timmermann, F. Bremmer, J. Waldthaler (2022). Visual perturbation suggests increased effort to maintain balance in early stages of Parkinson’s to be an effect of age rather than disease. Front Hum Neurosci 16:762380. doi: 10.3389/fnhum.2022.762380
- M. De Vitis, R. Breveglieri, K. Hadjidimitrakis, W. Vanduffel, C. Galletti, P. Fattori. The neglected medial part of macaque area PE: segregated processing of reach depth and direction. Brain Structure & Function (2019), 224(7), pp. 2537 – 2557. doi: 10.1007/s00429-019-01923-8
- T. L. Watson, M. Lappe. Fixation related shifts of perceptual localization counter to saccade direction. Journal of Vision 2019;19(13):18. doi: 10.1167/19.13.18
